Медицинская реабилитация

Физиология центральной нервной системы. Смирнов В.М. и др.
Физиология центральной нервной системы. Смирнов В.М. и др. В учебном пособии рассмотрены механизмы деятельности нейронов и их ансамблей, механизмы возбуждения и торможения нейронов центральной нервной системы (ЦНС), проведения возбуждения по нервным волокнам и синапсам; свойства нервных центров, функции различных отделов ЦНС; механизмы и принципы управления функциями органов и систем организма; координационная и интегрирующая роль ЦНС; общие принципы организации поведения организма и частные поведенческие реакции. Для студентов медицинских высших учебных заведений. Может быть использовано студентами психологических факультетов вузов.
Оглавление
5.1. Структурно-функциональная характеристика
5.2. Проводниковая функция
5.3. Рефлексы спинного мозга
5.4. Механизм шагательного рефлекса
5.5. Регуляция тонуса мышц
5.1. Структурно-функциональная характеристика
Сегменты и утолщения. Спинной мозг представляет собой тяж длиной около 45 см у мужчин и около 42 см — у женщин, имеет сегментарное строение (31 — 33 сегмента); каждый его сегмент связан с определенной частью тела. Спинной мозг включает пять отделов: шейный (C1 —С8), грудной (Th1 — Th12), поясничный (L1 — L5), крестцовый (S1 —S5) и копчиковый —Со3).
В процессе эволюции сформировалось два утолщения: шейное (сегменты, иннервирующие верхние конечности) и пояснично-крестцовое (сегменты, иннервирующие нижние конечности) как результат повышенной нагрузки на эти отделы спинного мозга. У некоторых видов животных подобных утолщений нет, например у змеи, которая передвигается благодаря равномерному участию в процессе движения всей мускулатуры тела. Тренировка любого органа обеспечивает прогрессивное его развитие не только в фило-, но и в онтогенезе, естественно, при этом совершенствуется и функция. Орган, который не получает достаточной нагрузки, постепенно атрофируется. В указанных утолщениях спинного мозга соматические нейроны наиболее крупные, их больше, в каждом корешке этих сегментов больше нервных волокон, они имеют наибольшую толщину.
Общее количество нейронов спинного мозга — около 13 млн (3% — мотонейроны, 97% — вставочные нейроны, относящиеся в том числе и к вегетативной нервной системе).
Классификация нейронов. Их целесообразно классифицировать по нескольким признакам: 1) по отделу нервной системы (нейроны соматической и вегетативной нервной системы); 2) по назначению, т.е. по направлению информации (эфферентные, афферентные, вставочные, ассоциативные); 3) по влиянию (возбуждающие и тормозные).
Эфферентные нейроны спинного мозга, относящиеся к соматической нервной системе, являются эффекторными, поскольку они иннервируют непосредственно рабочие органы — эффекторы (скелетные мышцы), их называют мотонейронами. Различают а- и у-мотонейроны. а-мотонейроны иннервируют экстрафузальные мышечные волокна (скелетная мускулатура), их аксоны характеризуются высокой скоростью проведения возбуждения — 70—120 м/с. а-мотонейроны подразделяют на две подгруппы: а1 — быстрые, иннервирующие быстрые белые мышечные волокна, их лабильность достигает 50 имп/с, и а2 — медленные, иннервирующие медленные красные мышечные волокна, их лабильность — 10 — 15 имп/с. Низкая лабильность а-мотонейронов объясняется длительной следовой гиперполяризацией, сопровождающей ПД. На одном а-мотонейроне насчитывается до 20 тыс. синапсов: от кожных рецепторов, проприорецепторов и нисходящих путей вышележащих отделов ЦНС. у- мотонейроны рассеяны среди а-мотонейронов, их активность регулируется нейронами вышележащих отделов ЦНС, они иннервируют интрафузальные мышечные волокна мышечного веретена (мышечного рецептора). При изменении сократительной деятельности интрафузальных волокон под влиянием Y-мотонейронов изменяется активность мышечных рецепторов. Импульсация от мышечных рецепторов активирует а-мотонейроны этой же мышцы и тормозит а-мотонейроны мышцы-антагониста, тем самым регулируется тонус скелетных мышц и двигательные реакции. Эти нейроны обладают высокой лабильностью — до 200 имп/с, но их аксонам свойственна более низкая скорость проведения возбуждения — 10 — 40 м/с.
Афферентные нейроны соматической нервной системы локализуются в спинальных ганглиях и ганглиях черепных нервов. Их отростки, проводящие афферентную импульсацию от мышечных, сухожильных и кожных рецепторов, вступают в соответствующие сегменты спинного мозга и образуют синаптические контакты либо непосредственно на а-мотонейронах (возбуждающие синапсы), либо на вставочных нейронах.
Вставочные (промежуточные) нейроны устанавливают связь с мотонейронами спинного мозга, с чувствительными нейронами, а также обеспечивают связь спинного мозга с ядрами ствола мозга, а через них — с корой большого мозга. Вставочные нейроны могут быть как возбуждающими, так и тормозными, имеющими высокую лабильность — до 1000 имп/с.
Нейроны симпатической нервной системы являются также вставочными, расположены в боковых рогах грудного, поясничного и частично шейного отделов спинного мозга (С8 —L2), они фоновоактивны, частота разрядов 3 — 5 имп/с. Нейроны парасимпатического отдела вегетативной нервной системы также вставочные, локализуются в сакральном отделе спинного мозга (S2— S4) и также фоновоактивны.
Ассоциативные нейроны образуют собственный аппарат спинного мозга, устанавливающий связь между сегментами и внутри сегментов. Ассоциативный аппарат спинного мозга участвует в координации позы, тонуса мышц, движений.
Ретикулярная формация спинного мозга состоит из тонких перекладин серого вещества, пересекающихся в различных направлениях, ее нейроны имеют большое количество отростков. Ретикулярная формация обнаруживается на уровне шейных сегментов между передними и задними рогами, а на уровне верхнегрудных сегментов — между боковыми и задними рогами в белом веществе, примыкающем к серому.
Нервные центры спинного мозга. Здесь находятся центры регуляции большинства внутренних органов и скелетной мускулатуры. Центры симпатического отдела вегетативной нервной системы локализованы в следующих сегментах: центр зрачкового рефлекса — С8—Th2, регуляции деятельности сердца — Th—Th5, слюноотделения — Th2—Th4, регуляции функции почек — Th5—L3; сегментарно расположены центры, регулирующие функции потовых желез и сосудов, гладких мышц внутренних органов, центры пиломоторных рефлексов.
Парасимпатическую иннервацию получают из спинного мозга (S2—S4) все органы малого таза: мочевой пузырь, часть толстой кишки ниже ее левого изгиба, половые органы. У мужчин парасимпатическая иннервация обеспечивает рефлекторный компонент эрекции, у женщин — сосудистые реакции клитора, влагалища.
Центры управления скелетной мускулатурой находятся во всех отделах спинного мозга и иннервируют по сегментарному принципу скелетную мускулатуру шеи (Ci—С4), диафрагмы (С3—С5), верхних конечностей (С5—Th2), туловища (Th3—Li) и нижних конечностей (L2—S5).
Повреждения определенных сегментов спинного мозга или его проводящих путей вызывают специфические двигательные нарушения и расстройства чувствительности.
Каждый сегмент спинного мозга участвует в чувствительной иннервации трех дерматомов. Имеется дублирование и двигательной иннервации скелетных мышц, что повышает надежность их деятельности.
Функции спинного мозга — проводниковая и рефлекторная.
5.2. Проводниковая функция
Проводниковая функция осуществляется с помощью нисходящих и восходящих путей.
Афферентная информация поступает в спинной мозг через задние корешки, эфферентная импульсация и регуляция функций различных органов и тканей организма осуществляется через передние корешки (закон Белла —Мажанди).
Каждый корешок представляет собой множество нервных волокон. Например, дорсальный корешок кошки включает 12 тыс., а вентральный — 6 тыс. нервных волокон.
Все афферентные входы в спинной мозг несут информацию от трех групп рецепторов: 1) от кожных рецепторов (болевых, температурных, прикосновения, давления, вибрации); 2) от проприорецепторов (мышечных — мышечных веретен, сухожильных — рецепторов Гольджи, надкостницы и оболочек суставов); 3) от рецепторов внутренних органов — висцерорецепторов (механои хеморецепторов).
Таблица 5.1
Основные проводящие пути спинного мозга
| Восходящие (чувствительные) пути | Физиологическое значение |
|---|---|
| Клиновидный пучок (пучок Бурдаха) проходит в задних столбах, импульсация поступает в кору | Осознаваемая проприорецептивная импульсация от нижней части туловища и ног |
| Тонкий пучок (пучок Голля), проходит в задних столбах, импульсация поступает в кору | Осознаваемая проприорецептивная импульсация от верхней части туловища и рук |
| Задний спинно-мозжечковый (Флексига) | Не осознаваемая проприорецептивная импульсация |
| Передний спинно-мозжечковый (Говерса) | Тоже |
| Латеральный спиноталамический | Болевая и температурная чувствительность |
| Передний спиноталамический | Тактильная чувствительность, прикосновение, давление |
| Нисходящие (двигательные) пути | Физиологическое значение |
| Латеральный кортикоспинальный (пирамидный) | Импульсы к скелетным мышцам, произвольные движения |
| Передний кортикоспинальный (пирамидный) | То же |
| Руброспинальный (Монакова), проходит в боковых столбах | Импульсы, поддерживающие тонус скелетных мышц |
| Ретикулоспинальный, проходит в передних столбах | Импульсы, поддерживающие тонус скелетных мышц с помощью возбуждающих и тормозящих влияний на аи у-мотонейроны, а также регулирующие состояние спинальных вегетативных центров |
| Вестибулоспинальный, проходит в передних столбах | Импульсы, обеспечивающие поддержание позы и равновесия тела |
| Тектоспинальный, проходит в передних столбах | Импульсы, обеспечивающие осуществление зрительных и слуховых двигательных рефлексов (рефлексов четверохолмия) |
Медиатором первичных афферентных нейронов, локализующихся в спинальных ганглиях, является, по-видимому, субстанция Р.
Значение афферентной импулъсации, поступающей в спинной мозг, заключается в следующем: 1) участие в координационной деятельности ЦНС по управлению скелетной мускулатурой; при выключении афферентной импульсации от рабочего органа управление им становится несовершенным; 2) участие в процессах регуляции функций внутренних органов; 3) поддерживает тонус ЦНС; при выключении афферентной импульсации наступает уменьшение суммарной тонической активности ЦНС; 4) несет информацию об изменениях окружающей среды. Основные проводящие пути спинного мозга приведены в табл. 5.1.
5.3. Рефлексы спинного мозга
В данном разделе рассматриваются только соматические рефлексы (вегетативные рефлексы см. в разд. 7.10).
Рефлексы спинного мозга достаточно просты. По форме это в основном сгибательные и разгибательные рефлексы сегментарного характера. Надсегментарные рефлексы наряду с сегментарными осуществляются только с помощью шейного отдела (см. далее подразд. 8.1.3). Сила и длительность спинальных рефлексов, как и рефлексов других отделов ЦНС, увеличиваются при повторном раздражении, при увеличении площади раздражаемой рефлексогенной зоны вследствие суммации возбуждения, а также при увеличении силы стимула.
Классификация соматических рефлексов спинного мозга (общие принципы). Все спинальные рефлексы можно объединить в две группы по следующим признакам. Во-первых, по рецепторам, раздражение которых вызывает рефлекс: а) проприорецептивные, б) висцерорецептивные и в) кожные рефлексы (защитные). Рефлексы, возникающие с проприорецепторов, участвуют в формировании акта ходьбы и регуляции мышечного тонуса. Висцерорецептивные рефлексы возникают с интерорецепторов (рецепторов внутренних органов) и проявляются в сокращениях мышц передней брюшной стенки, грудной клетки и разгибателей спины.
Во-вторых, спинальные рефлексы целесообразно объединить по органам (эффекторам рефлекса): а) рефлексы конечностей; б) брюшные; в) яичковый; г) анальный. Рассмотрим каждую группу этих рефлексов.
Рефлексы конечностей. Данная группа рефлексов наиболее обширная, в клинической практике их исследуют наиболее часто. Однако отдельные рефлексы описывают без взаимной связи. Если в качестве объединяющего признака рефлексов конечностей принять характер ответной реакции, то все их можно объединить в четыре группы: 1) сгибательные; 2) разгибательные; 3) ритмические; 4) познотонические.
Сгибательныерефлексы делятся на фазные и тонические.
Фазные рефлексы — это однократное сгибание конечности при однократном раздражении кожи или проприорецепторов. Одновременно с возбуждением мотонейронов мышц-сгибателей происходит реципрокное торможение мотонейронов мышц- разгибателей. Рефлексы, возникающие с рецепторов кожи, являются полисинаптическими, они имеют защитное значение (рис. 5.1, а). Например, погружение в слабый раствор серной кислоты лапки спинальной лягушки, подвешенной на крючке, или щипок кожи конечности пинцетом вызывают отдергивание конечности вследствие сгибания ее в коленном суставе, а при более сильном раздражении — и в тазобедренном суставе. Рефлексы, возникающие с проприорецепторов, могут быть моносинаптическими и полисинаптическими, например шейные познотонические рефлексы. Фазные рефлексы с проприорецепторов участвуют в формировании акта ходьбы. По степени выраженности фазных сгибательных и разгибательных рефлексов определяют состояние возбудимости ЦНС и возможные ее нарушения.
Тонические сгибательные, а также разгибательные рефлексы возникают при длительном растяжении мышц, их главное назначение — поддержание позы. Тоническое сокращение скелетных мышц является фоновым для осуществления всех двигательных актов, осуществляемых с помощью фазических сокращений мышц.
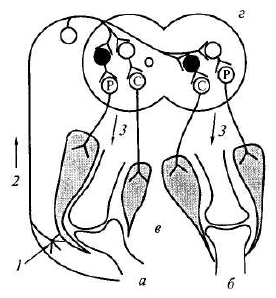
Рис. 5.1. Рефлексы нижних конечностей:
а — дуга сгибательного (защитного) рефлекса; б — дуга перекрестного разгибательного рефлекса; в — коленные суставы с мышцами; г — сегмент спинного мозга; 1 — раздражение кожных рецепторов; 2 — афферентный путь (—); 3 — эфферентные пути (I) от амотонейронов центров сгибания (С) и разгибания (Р). Вставочные нейроны: ф < тормозные, О—< возбуждающие
В клинике исследуют несколько сгибательных фазных рефлексов: локтевой и ахиллов (проприоцептивные рефлексы), подошвенный (кожный рефлекс). Локтевой рефлекс выражается в сгибании руки в локтевом суставе, возникает при ударе молоточком по сухожилию m. biceps brachii (при вызове рефлекса рука должна быть слегка согнута в локтевом суставе), его дуга замыкается в пятом-шестом шейных сегментах спинного мозга (С5—С6). Ахиллов рефлекс выражается в подошвенном сгибании стопы в результате сокращения трехглавой мышцы голени, возникает при ударе молоточком по ахиллову сухожилию, рефлекторная дуга замыкается на уровне крестцовых сегментов S1—S2. Подошвенный рефлекс — сгибание стопы и пальцев при штриховом раздражении подошвы, дуга рефлекса замыкается также на уровне S1—S2.
Разгибателъные рефлексы, как и сгибательные, бывают фазными и тоническими, возникают с проприорецепторов мышц-разгибателей, являются моносинаптическими. Одновременно со сгибательным рефлексом возникает перекрестный разгибательный рефлекс другой конечности (рис. 5.1, б).
Фазные рефлексы возникают в ответ на однократное раздражение мышечных рецепторов, например при ударе по сухожилию четырехглавой мышцы ниже надколенной чашечки. При этом возникает коленный разгибательный рефлекс вследствие сокращения четырехглавой мышцы (мотонейроны мышц-сгибателей во время разгибательного рефлекса тормозятся — постсинаптическое реципрокное торможение с помощью вставочных тормозных клеток Реншоу). Рефлекторная дуга коленного рефлекса замыкается во втором — четвертом поясничных сегментах (L2—L4). Фазные разгибательные рефлексы участвуют, как и сгибательные, в формировании акта ходьбы.
Тонические разгибательные рефлексы представляют собой длительное сокращение мышц-разгибателей при длительном растяжении их сухожилий. Их роль — поддержание позы. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикальной естественной позы. Тоническое сокращение мышц спины удерживает туловище в вертикальном положении, обеспечивая осанку человека. Тонические рефлексы на растяжение мышц (сгибателей и разгибателей) называют также миотатическими.
Рефлексы позы — перераспределение мышечного тонуса, возникающее при изменении положения тела или отдельных его частей. Рефлексы позы осуществляются с участием различных отделов ЦНС. На уровне спинного мозга замыкаются шейные позные рефлексы, наличие которых установил голландский физиолог Р. Магнус (1924) в специальных опытах на кошке. Имеется две группы этих рефлексов — возникающие при наклоне и при поворотах головы.
Первая группа рефлексов возникает при наклоне головы вниз (кпереди): увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц разгибателей задних конечностей, в результате чего передние конечности сгибаются, а задние разгибаются. При наклоне головы вверх (кзади) возникают противоположные реакции — передние конечности разгибаются вследствие увеличения тонуса их мышц- разгибателей, а задние конечности сгибаются вследствие повышения тонуса их мышц-сгибателей (рис. 5.2). Эти рефлексы возникают с проприорецепторов мышц шеи и фасций, покрывающих шейный отдел позвоночника. В условиях естественного поведения они увеличивают животному шансы достать пищу, находящуюся выше или ниже головы.
Рефлексы позы верхних конечностей у человека, по-видимому, утрачены, рефлексы нижних конечностей выражаются не в сгибании или разгибании, а в перераспределении мышечного тонуса, обеспечивающего сохранение естественной позы.
Вторая группа шейных позных рефлексов возникает с тех же рецепторов, но только при поворотах головы вправо или влево.
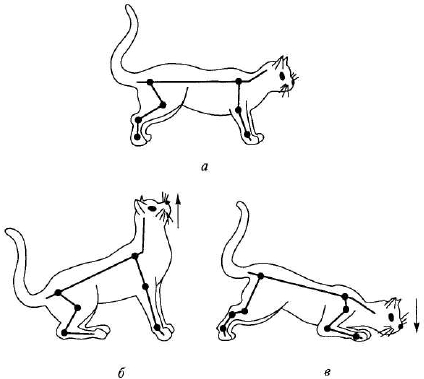
При а — до изменения положения головы; б — при пассивном поднимании (Т), в — при пассивном опускании (X) головы этом повышается тонус мышц-разгибателей обеих конечностей на стороне, куда повернута голова, и повышается тонус мышц-сгибателей на противоположной стороне. Рефлекс направлен на сохранение позы, которая может быть нарушена вследствие изменения положения центра тяжести после поворота головы. Центр тяжести смещается в сторону поворота головы — именно на этой стороне повышается тонус мышц- разгибателей обеих конечностей. Подобные рефлексы наблюдаются и у человека.
Ритмические рефлексы — многократное повторное сгибание и разгибание конечностей. Примерами этих рефлексов могут быть рефлексы потирания у лягушки, чесательный и шагательный рефлексы у собаки. Рефлекс потирания заключается в том, что после смазывания раствором кислоты кожи бедра спинальная лягушка (лягушка, у которой удален головной мозг) многократно потирает этот участок, т.е. пытается освободиться от раздражителя. Слабое раздражение кожи туловища у собаки вызывает почесывание этого участка задней конечностью. Чесательный рефлекс — аналог рефлекса потирания у лягушки. Шагательный рефлекс наблюдают у спинальной собаки, подвешенной с помощью лямок в станке.
Брюшные рефлексы (верхний, средний и нижний) проявляются при штриховом раздражении кожи живота; выражаются в сокращении соответствующих участков мускулатуры стенки живота. Это защитные рефлексы. Для вызова верхнего брюшного рефлекса раздражение наносят параллельно нижним ребрам непосредственно под ними, дуга рефлекса замыкается на уровне грудных сегментов спинного мозга (Th8—Th9). Средний брюшной рефлекс вызывают раздражением на уровне пупка (горизонтально), дуга рефлекса замыкается на уровне Th9—Th10. Для получения нижнего брюшного рефлекса раздражение наносят параллельно паховой складке (рядом с ней), дуга рефлекса замыкается на уровне Th11—Th12.
Кремастерный (яичковый рефлекс) заключается в сокращении m. cremaster и поднимании мошонки в ответ на штриховое раздражение верхней внутренней поверхности кожи бедра (кожный рефлекс), это также защитный рефлекс; его дуга замыкается на уровне L1—L2.
Анальный рефлекс выражается в сокращении наружного сфинктера прямой кишки в ответ на штриховое раздражение или укол кожи вблизи заднего прохода, дуга рефлекса замыкается на уровне S2 — S5.
5.4. Механизм шагательного рефлекса
Способ вызова рефлекса. Рефлекс вызывается однократным раздражением кожи конечности. Он выражается в сгибании этой конечности с одновременным дополнительным разгибанием противоположной задней конечности — перекрестный разгибательный рефлекс (см. рис. 5.1, б). Затем согнутая конечность разгибается, опускается вниз, разогнутая — сгибается и поднимается вверх и т.д. Конечности опоры не касаются (собака зафиксирована в станке с помощью лямок) и без дополнительного раздражения продолжают ритмично поочередно сгибаться и разгибаться, т.е. «шагать» в воздухе. Рефлекс может продолжаться часами, так как устранено влияние коры большого мозга. При неповрежденных сегментах, иннервирующих передние конечности (С5—Th2), и более длительном раздражении задней конечности в рефлекс вовлекаются также и передние конечности. Это наблюдение показывает наличие межсегментарной связи на уровне спинного мозга, играющей важную роль в ритмических движениях конечностей. Поочередное сокращение и расслабление мышц-сгибателей и разгибателей осуществляется в результате взаимодействия процессов возбуждения и торможения в соответствующих центрах спинного мозга под влиянием импульсов, поступающих в мозг от проприорецепторов.
Роль проприорецепторов в осуществлении шагательного рефлекса определяется их расположением.
Мышечные веретена (мышечные рецепторы) расположены параллельно скелетной мышце: своими концами крепятся к соединительно-тканной оболочке (перимизию) пучка экстрафузальных мышечных волокон при помощи напоминающих сухожилия полосок соединительной ткани длиной 0,5 — 1 мм. Поэтому при расслаблении (удлинении) мышцы растягиваются и мышечные рецепторы, что и ведет к их возбуждению.
Мышечный рецептор состоит из нескольких поперечнополосатых интрафузальных мышечных волокон (от лат. fusus — веретено), окруженных соединительнотканной капсулой. Длина интрафузальных мышечных волокон равна 4-7 мм, толщина — 15-30 мкм. Длина экстрафузальных мышечных волокон — от нескольких миллиметров до многих сантиметров, толщина — 50-100 мкм. Вокруг средней части мышечного веретена несколько раз обвивается окончание одного афферентного волокна. Многие мышечные веретена иннервируются и другим (более тонким) афферентным волокном, которое также обвивает в виде спирали мышечный рецептор, но периферические его участки. Импульсы от мышечных рецепторов возбуждают нейроны своего центра и тормозят нейроны центраантагониста. Ближе к концам мышечного веретена подходят двигательные нервные окончания, являющиеся аксонами -мотонейронов спинного мозга. Их импульсация вызывает сокращение интрафузальных мышечных волокон (мышечных волокон рецептора).
Сухожильные рецепторы (рецепторы Гольджи) заключены в соединительно-тканную капсулу и локализуются в сухожилиях скелетных мышц вблизи от сухожильно-мышечного соединения. Рецепторы представляют собой безмиелиновые окончания толстого миелинового афферентного волокна (подойдя к капсуле рецепторов Гольджи, это волокно теряет миелиновую оболочку и делится на несколько окончаний). Сухожильные рецепторы крепятся относительно скелетной мышцы последовательно, что обеспечивает их раздражение при натяжении сухожилия. Поэтому сухожильные рецепторы посылают информацию в мозг о том, что мышца сокращена (напряжено и сухожилие), а мышечные рецепторы — о том, что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мышц-сгибателей это возбуждение выражено слабее, чем у разгибателей).
Элемент шагательного рефлекса — поочередное сокращение и расслабление скелетной мышцы под влиянием импульсов от проприорецепторов, поступающих в свой центр.
Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от них поступают к своим а-мотонейронам спинного мозга и возбуждают их (рис. 5.3, а). Далее а-мотонейроны посылают импульсы к этой же скелетной мышце, что ведет к ее сокращению. Как только мышца сократилась (рис. 5.3, б), возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), начинают возбуждаться сухожильные рецепторы. Импульсы от последних поступают тоже в первую очередь в свой центр в спинном мозге, но к тормозным клеткам Реншоу. Возбуждение тормозных клеток вызывает торможение амотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабление (удлинение) ведет снова к возбуждению мышечных веретен и а-мотонейронов — мышца вновь сокращается. Вследствие ее сокращения снова возбуждаются сухожильные рецепторы и тормозные клетки в спинном мозге, что вновь ведет к расслаблению скелетной мышцы и т.д. Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце- сгибателю, и к мышце-разгибателю. При этом расслабление скелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу.
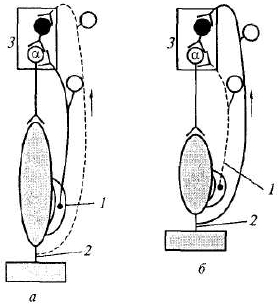
Для обеспечения поочередного сгибания и разгибания конечностей при шагательном рефлексе мышцы-сгибатели и мышцы-разгибатели должны сокращаться и расслабляться последовательно друг за другом, что достигается с помощью торможения центра-антагониста при возбуждении центра-агониста. Причем если на одной ноге сокращены мышцы-сгибатели, на другой ноге должны сокращаться мышцы- разгибатели, что обеспечивается поступлением афферентных импульсов от мышечных и сухожильных рецепторов и поочередным возбуждением и торможением центров сгибателей и разгибателей. На одноименной же стороне при возбуждении центра мышцы-сгибателя тормозится центр мышцы-разгибателя (см. рис. 5.1).
а Шагательные координированные движения у спинального животного возможны в отсутствие обратной афферентации от проприорецепторов. Они осуществляются с помощью межсегментарных связей на уровне спинного мозга. О наличии межсегментарных связей свидетельствует также факт вовлечения в шагательный рефлекс всех четырех конечностей спинальной собаки при достаточно длительном и сильном раздражении одной конечности.
5.5. Регуляция тонуса мышц
Тонус мышц исследуют после поперечной перерезки спинного мозга. Сразу после перерезки в эксперименте или повреждения спинного мозга у человека в случае травмы наблюдаются мышечная атония и отсутствие рефлексов (спинальный шок). Главной причиной спинального шока является выключение влияния на спинной мозг вышележащих отделов ЦНС (повторная перерезка спинного мозга ниже первой повторно шок не вызывает). Спинальный шок у лягушек длится несколько минут, у собак — несколько дней, у человека — около 2 мес. После исчезновения спинального шока тонус мышц, иннервируемых посредством сегментов спинного мозга, которые находятся ниже перерезки (повреждения), резко повышается. Сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения спинного мозга усиливаются. Механизм повышения тонуса мышц нижних конечностей в случае повреждения спинного мозга (после окончания спинального шока) отражает схема, представленная на рис. 5.4.
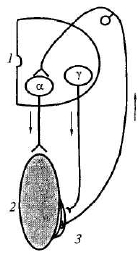
Рис. 5.4. Схема механизма гипертонуса мышц нижних конечностей в случае повреждения спинного мозга: 1 — полусегмент спинного мозга; 2 — скелетная мышца; 3 — мышечное веретено; Т — афферентный путь; X — эфферентные пути; а-мотонейрон, иннервирующий скелетную мышцу; у-мотонейрон, иннервирующий интрафузальные мышечные волокна (проприорецептора)
Гипертонус имеет рефлекторную природу, он развивается вследствие афферентной импульсации от мышечных рецепторов. Деафферентация, например, нижней конечности у лягушки ведет к исчезновению у нее тонуса (опыт Бронжеста).
Имеется два фактора, обеспечивающих афферентную импульсацию от мышечных рецепторов по задним корешкам спинного мозга, которая ведет к возбуждению а-мотонейронов спинного мозга и сокращению скелетных мышц при повреждении спинного мозга у человека: 1) спонтанная активность мышечных рецепторов; 2) спонтанная активность у-мотонейронов, которая реализуется следующим образом. Возбуждение у-мотонейронов вызывает возбуждение и сокращение иннервируемых ими интрафузальных мышечных волокон, в результате чего увеличивается натяжение мышечного рецептора, поскольку концы его зафиксированы на скелетной мышце. Вследствие натяжения мышечного веретена раздражаются и возбуждаются мышечные рецепторы, обеспечивающие импульсацию к а-мотонейронам и возбуждение их. В свою очередь а-мотонейроны посылают импульсы к скелетной мышце и вызывают ее постоянное (тоническое) сокращение.
Мы рассмотрели рефлекторную и проводниковую функции изолированного спинного мозга. В нормальных условиях деятельность спинного мозга контролируется вышележащими отделами ЦНС посредством импульсации ко всем его нервным элементам.