Медицинская реабилитация
1.7. Нейрофизиологическая характеристика двигательной единицы и организация спинальной сегментарной деятельности
Двигательные нервные клетки, или мотонейроны, являются выходными (эфферентными) клетками спинного мозга, осуществляющими передачу выработанных в спинном мозге сигналов к скелетным мышцам. Мотонейроны и периферические нервы, состоящие из их аксонов, оказывают нейротрофическое влияние на иннервированные ими мышцы. Установлено, что их воздействие на мышцу не зависит от возбуждающих двигательных импульсов. Можно частично пережать нерв, блокируя импульсы, но не нарушая непрерывность аксоплазмы. В этом случае изменения, характерные для денервации мышцы (атрофия, повышение возбудимости и др.), не наступают. Изменения, возникающие в мышце после денервации, принято считать следствием утраты какого-то трофического вещества, переходящего в области концевой пластинки из нервного окончания в мышечное волокно и снижающего возбудимость мембраны при действии ацетилхоли-на вне этой области.
В настоящее время под трофическим веществом подразумеваются компоненты быстрого ортоградного аксонального транспорта: аминокислоты, сахара, жирные кислоты, липиды, белки и даже целые органеллы. Эти составные части либо доставляются в аксолемму по всей длине нервных волокон, либо направляются к нервным окончаниям, где они включаются в пресинаптические плазматические мембраны и синаптические пузырьки.
Иннервация, типичная для позвоночных, характеризуется наличием некоторого числа отдельных мышечных волокон, возбуждаемых ветвями одного двигательного нервного волокна. Каждый мотонейрон вместе с его аксоном и иннервируемыми им мышечными волокнами образует двигательную единицу. Мышечные волокна одной двигательной единицы функционируют как единое целое, поэтому при возбуждении двигательного нерва они сокращаются по принципу «все или ничего». Зона двигательной единицы в поперечнике имеет круглую форму и обычно перекрывается зонами двух или трех соседних. В зависимости от функциональных характеристик различные мышцы имеют разные размеры двигательных единиц, то есть число мышечных волокон в ней может колебаться от нескольких единиц (например, в мышцах глазного яблока) до нескольких сотен (в тонических мышцах конечностей). В мелких мышцах двигательные единицы могут включать 10—25 мышечных волокон, тогда как в больших мышцах, несущих познотоническую функцию, это число может доходить до 2000 (Л. О. Бадалян, И. А. Скворцов, 1986). Как правило, двигательные единицы, контролирующие мелкие движения, состоят из меньшего числа мышечных волокон, что обеспечивает более тонкую градацию, то есть более тонкую регуляцию мышцы.
Количество мышечных волокон, которые относятся к одному двигательному нейрону, прямо пропорционально его размеру и обратно пропорционально степени его возбудимости. Поэтому с функциональной точки зрения двигательные мотонейроны разделяют на две группы: медленные и быстрые. Медленные двигательные единицы включают медленный низкопороговый мотонейрон, который иннервирует однотипные медленные мышечные волокна; быстрые двигательные единицы — соответственно быстрые высокопороговые мотонейроны и быстрые мышечные волокна. В пределах одной мышцы двигательные единицы, занимающие меньшую зону, являются медленными и, наоборот, более крупные двигательные единицы — быстрыми. При электромиографии установлено, что низкопороговые двигательные единицы вызывают сокращение мышцы с частотой 4—5 Гц, которое повышается до 6—10 Гц перед активизацией потенциалов действия двигательных единиц со следующим (более высоким) порогом. Высокопороговые потенциалы действия двигательных единиц разряжаются с частотой 30—50 Гц. Все мышечные волокна одной двигательной единицы имеют одинаковый гистохимический тип.
Способность мышцы к механической деятельности, оцениваемую по силе, скорости сокращения, расслабления и другим функциональным показателям, определяют как ее сократимость. Величина сокращения (напряжения) мышцы зависит от частоты нервных импульсов в двигательном нерве, числа возбужденных нейромоторных (двигательных) единиц, соотношения между количеством участвующих в сокращении быстрых (фазических) и медленных (тонических) волокон, а также от механических свойств соединительной ткани и самих сократительных волокон. Нарастающее и длительное сокращение мышцы обеспечивается асинхронным возбуждением отдельных нейромоторных единиц с разными зонами. Скорость сокращения и расслабления мышцы связана с количеством быстрых и медленных волокон, составляющих нейромоторную единицу. Быстрее сокращаются и расслабляются мышцы, имеющие в составе нейромо-торные единицы с небольшим числом волокон.
Нервные волокна нейронов спинного мозга классифицируют по скорости проведения возбуждения и толщине (диаметру) на группы А, В, С (см. табл. 1). Нервные волокна группы А проводят импульсы со скоростью до 120 м/с. Они имеют большой диаметр поперечного сечения и выраженную миелинизацию. Волокна группы В переносят импульсы со скоростью проведения 3—14 м/с и имеют малый поперечный размер и миелинизацию. Немиелинизиро-ванные волокна группы С обладают наименьшими скоростью проведения и диаметром. Миелиновый слой волокон обусловливает быстрое сальтаторное проведение импульсов. Наиболее скорые нервные волокна существуют для тех функций, которые требуют быстроты. Толстые миелинизированные, быстропроводящие волокна группы А подразделяют на 4 подгруппы — a, b, g, d (в порядке убывания скорости проведения импульса). Аксоны мотонейронов имеют высокую скорость проведения импульса возбуждения и относятся к волокнам группы А. Иннервацию фазных мышечных волокон осуществляют аксоны диаметром до 20 мкм и скоростью проведения импульса от 60 до 120 м/с. Тонические мышечные волокна иннервированы тонкими (диаметром до 10 мкм) аксонами со скоростью проведения импульса до 30 м/с, образующими множественные гроздьевидные окончания.
При возбуждении расположенных на соме и дендритах мотонейрона синаптических окончаний афферентных клеток или интернейронов (промежуточных, или вставочных) в мотонейроне под действием выделяемых окончаниями медиаторов возникает локальная деполяризация мембран — возбуждающий постсинаптический потенциал (ВПСП). При суммации ВПСП до критического уровня возникает потенциал действия (ПД), который состоит из нескольких различающихся по скорости развития компонентов (ПД сомы и дендритов, ПД начального сегмента аксона). Максимальная частота воспроизведения ПД в мотонейроне не превышает 200—300 импульсов в 1 с. Часто она существенно ниже (несколько десятков импульсов в 1 с) в связи с появлением вслед за ПД длительной следовой гиперполяризации (70— 170 мс). Следовую гиперполяризацию не следует смешивать с гиперполяризацией, вызванной работой клеток Реншо. По выраженности следовой гиперполяризации и частоте импульсации мотонейроны разделяют на две группы: фазические (быстрые) и тонические (медленные), которые иннервируют соответственно быстрые («белые») и медленные («красные») мышцы.
Основой торможения мотонейронов является появление в них при соответствующих синаптических влияниях локальной гиперполяризации мембран — тормозящего постсинаптического потенциала (ТПСП) примерно такой же длительности, как ВПСП.
ТПСП создаются интернейронами (клетки Реншо), синтезирующими и выделяющими соответственный тормозной медиатор, роль которого играют глицин и гамма-аминомасляная кислота.
Афферентные волокна отдают коллатерали, которые ветвятся и образуют синапсы на тормозных (вставочных) нейронах в промежуточной части спинного мозга. Тормозные нейроны в свою очередь посылают волокна к антагонистическим мотонейронам переднего рога, вызывая ТПСП в постсинаптической мембране мотонейрона мышцы-антагониста. ВПСП возникает в мембране мотонейрона после синаптического возбуждения и представляет собой явление деполяризации. ТПСП имеет всегда больший латентный период, чем возбуждающий, при ортодромном раздражении, что связано с наличием дополнительной синаптической задержки на вставочном тормозном нейроне.
Мотонейроны в спинном мозге собраны в группы (ядра) по функциональному признаку — иннервации определенных групп мышц. При возникновении рефлекторной двигательной реакции возбуждается сравнительно небольшое количество клеток ядра и соответственно этому активизируется лишь часть двигательных единиц. Остальные клетки остаются в состоянии подпорогового возбуждения, образуя «подпороговую кайму». При конвергенции к двигательному ядру двух и более афферентных волн локальные процессы суммируются и вовлекают в разряд большее количество мотонейронов, в результате чего возникает так называемое облегчение рефлекторного ответа. Способность переходить из подпорогового состояния в состояние активности у различных мотонейронов неодинакова. Клетки большего диаметра обладают более низкой возбудимостью (высоким порогом), чем клетки меньшего диаметра. При тормозящих влияниях наблюдается противоположный процесс — большее количество двигательных нервных клеток переходит в состояние подпорогового синаптического возбуждения и таким образом увеличивается «подпороговая кайма».
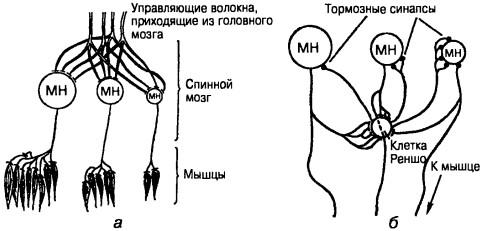
Рис. 15. Работа системы мотонейронов, управляющих сокращением мышцы: а — «принцип величины» в работе мотонейрон-ного пула, б — клетки Реншо предохраняют маленькие мотонейроны от перегрузки
Каждой крупной мышцей управляет своя группа мотонейронов — мотонейронный пул. Для управления мышцей не требуется командовать каждым мышечным волокном в отдельности. Достаточно изменять только силу сигнала, приходящего на пул, а нужный порядок включения мышечных волокон обеспечивается «геометрией» мотонейронов пула — их размерами и «топографией», то есть расположением аксонов в мышце. Мотонейроны отличаются размерами, причем чем крупнее нейрон, тем больше у него аксонных окончаний и тем больше число мышечных волокон он возбуждает. Самые крупные мотонейроны имеют окончания именно на тех мышечных волокнах, которые нужны для «авральной» работы — кратковременной, но с развитием больших усилий (рис. 15). Управляющие команды от верхних отделов головного мозга приходят по волокнам, которые равномерно распределены по всему мотонейронному пулу (например, каждое волокно, приходящее сверху, дает по 5 синапсов на каждом нейроне). Если при слабых входных сигналах возбуждено небольшое число управляющих волокон или по ним приходят импульсы с низкой частотой, возбудятся только самые мелкие мотонейроны, и мышца будет развивать небольшое усилие. С увеличением входного сигнала чем большее число управляющих волокон будет возбуждено, и большее число синапсов на мотонейроне будет активизировано, тем более крупные мотонейроны включатся в работу. Таким образом, управляя только одним параметром — числом возбужденных нисходя-
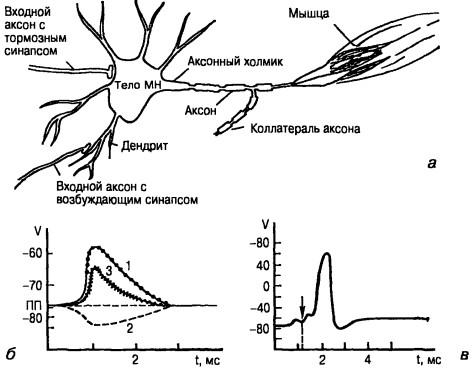
Рис. 16. Схематическое изображение мотонейрона кошки (а) и изменение потенциала в его теле при разных воздействиях (б), временное суммирование потенциалов. Первый синаптический потенциал был поДпороговым, но второй такой же потенциал (момент его возникновения отмечен стрелкой) сложился с первым и привел к возникновению импульса (в):
1 — ВПСП, возникающий в теле нейрона под действием нескольких синапсов, 2 — ТПСП, 3 — суммирование ВПСП и ТПСП
щих волокон, головной мозг включает в работу те мышечные волокна, которые нужны для выполнения требуемого движения (М. Б. Беркинблат, Е. Г. Глаголева, 1988).
Впервые гипотезу о том, что для управления мотонейронным пулом мышцы природа использует именно геометрические различия размеров тел нервных клеток, высказал в 1965 г. американский ученый Ханнеман. В дальнейшем эта гипотеза была подтверждена разнообразными экспериментами и получила название «принцип величины». Геометрический подход состоит в том, что свойства возбудимых клеток и тканей, а значит, и выполняемые ими функции во многом определяются их геометрической структурой: формой клеток, соотношением их размеров, взаимным
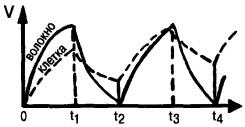
Рис. 17. Нарастание и спад потенциала в нервной клетке и в волокне при пропускании импульсов тока (ток включен в момент времени 0 и выключен в момент tj, второй импульс тока начинается в момент t2 и заканчивается в момент t3); у клетки потенциал нарастает и падает медленнее, чем у волокна; у нее сдвиг потенциала в ответ на второй импульс суммируется с ответом на первый импульс
расположением и связями. В настоящее время геометрический подход в электрофизиологии широко распространен и является вполне естественным: ведь поведение, например, нейрона существенно зависит от распределения потенциала на его мембране и от токов, которые текут в клетке и окружающей ее среде. Но токи и напряжения, в свою очередь, зависят от распределения сопротивления и емкостей, а это распределение определяется формой клетки. В ходе расчетов оказалось, что постсинаптический потенциал, создаваемый одинаковыми синапсами в разных нервных клетках, имеющих сферическую поверхность, будет прямо пропорционален их входному сопротивлению, которое, в свою очередь, обратно пропорционально квадрату диаметра сферической клетки. Следовательно, при одинаковом воздействии постсинаптический потенциал в клетке диаметром 10 мкм будет в 100 раз больше, чем в клетке диаметром в 100 мкм. По этому поводу известный русский ученый М. М. Бонгард сказал: «Это понятно. Спичкой можно зажечь тонкую хворостинку, но нельзя зажечь бревно».
Примерно одновременное воздействие многих синапсов, находящихся на разных участках клеточной мембраны, суммируется (пространственная суммация). Однако, как правило, синапсы срабатывают неодновременно, то есть фазы их не совпадают. В таком случае процесс называют пространственно-временной суммацией (рис. 16). Для этих двух видов суммации большое значение имеет, насколько долго сохраняется след от синаптического воздействия, что зависит, в свою очередь, от емкостных свойств нейрона (рис. 17). Если возникший потенциал спадает достаточно медленно (как в нервной клетке), то действие второго
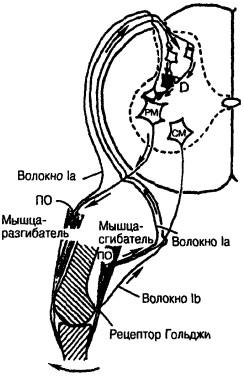
Рис. 18. Пресинаптическое торможение. Волокна 1а от первичных окончаний (ПО) веретен мышцы-сгибателя и мышцы-разгибателя оканчиваются на клетке заднего рога, образующей синапсы на D-клетке (СМ). Последняя образует синапсы на окончаниях афферентных волокон, в данном случае на афферентах разгибательно-го мотонейрона (РМ) и вызывает торможение. Волокна от сухожильного рецептора также могут оканчиваться на D-клетке и тормозить афференты веретен
синапса, включившегося по-_зднее, суммируется с остаточным потенциалом от первого. В противном случае (в кабелях-аксонах) суммируется только действие тех синапсов, которые сработали практически одновременно.
Способ управления мотонейронным пулом, построенный на основе геометрического подхода, имеет один недостаток: сигнал, необходимый для включения в работу больших мотонейронов, слишком велик для маленьких. Он заставил бы их работать со слишком большой частотой, что может привести к гибели этих клеток. Чтобы этого не произошло, в мотонейронном пуле есть специальное защитное устройство — клетки Реншо. К последним идут специальные отростки аксонов мотонейронов, причем в основном от больших мотонейронов. Аксоны тормозных клеток Реншо (D-клетки) оканчиваются преимущественно на маленьких мотонейронах (рис. 18). Если сигнал, приходящий сверху, велик, то к маленьким мотонейронам приходит, с одной стороны, этот слишком большой для них сигнал, а с другой — тормозной сигнал от клеток Реншо. Противоположные по действию сигналы складываются, и маленькие мотонейроны работают в нормальном режиме. Таким образом, возвратное торможение локализует рефлекторные ответы, устраняя возбуждение в окружающих популяциях моонейронов. Поэтому, если отводить электромиограмму от мышц, в которых поддерживается фоновая рефлекторная активность, то в течение 50— ЮОмс после миотатического рефлекса в таких мышцах наблюдается период молчания. Причем функция этого специализированного торможения проявляется в разгибательных и сгибательных мышцах. Длительное снижение возбудимости клетками Реншо при аутогенном торможении и в период молчания сильнее выражено в тонических мотонейронах, хотя резкой грани между тоническими и фазическими мотонейронами не существует. Более выраженное торможение тонических мотонейронов способствует подавлению тонуса мышцы и снижению ее сопротивления фазическим ответам.
Кроме торможения, следует учитывать и возвратное облегчение, хотя оно значительно слабее, чем тормозное действие.
Пресинаптическое торможение — важная система с отрицательной обратной связью, позволяющая выключить афферентные импульсы при их поступлении в ЦНС. Благодаря этому механизму подавляются сигналы, не имеющие значения, и пропускаются только действительно важные. Афферентные волокна или их коллатерали оканчиваются на определенных вставочных нейронах дорсального рога. Они, в свою очередь, образуют синапсы на афферентных волокнах и тормозят их разряды. Аксоны, подходящие к афферентным окончаниям, деполяризуют последние и таким образом уменьшают количество выделяемого медиатора. Существует две системы пресинаптического торможения. Первую составляют кожные и мышечные афференты с высоким порогом, оканчивающиеся на вставочных нейронах, а затем, через D-клетки, на других афферентах. В эту систему входят рецепторы движения волосков, прикосновения и давления, мышечные рецепторы давления и, возможно, болевые волокна, вызывающие сгибательный рефлекс. Сгибательный рефлекс является одним из проявлений действия этой системы. Поэтому вся система называется афферентами сгибательного рефлекса. Другая система пресинаптического торможения связана с мышечными афферентами группы I, действующими при проприоцептивных реакциях мышцы. В эту группу входят афференты 1а и lb. При краткой тетанизации афферентов торможение длится несколько сотен миллисекунд (см. рис. 18).
Таким образом, аксонные коллатерали и клетки Реншо образуют системы отрицательной обратной связи, которые влияют на мото- и интернейроны. В результате действия такой системы моторные ядра могут активно удерживаться в состоянии низкой активности, достаточной для осуществления обычной двигательной деятельности и сохраняющей в то же время большие резервные возможности на случай резкого повышения требований к мышечной активности.
Возбуждение двигательного нейрона ведет к возникновению волны деполяризации, которая распространяется по аксону со скоростью 30—80 м/с. Как только волна деполяризации достигнет концевых нервных разветвлений, освобождается химический медиатор ацетилхолин, который проходит через синаптическую щель за 10—50 мкс. Ацетилхолин воспринимается рецепторами, которые располагаются на постсинаптической концевой пластинке, обычно на середине мышечного волокна. Если воспринято достаточно ацетилхолина, то концевая пластинка деполяризуется и волна деполяризации распространяется по мышечному волокну в обоих направлениях со скоростью от 1,5 до 6,5 м/с. Восстановление состояния покоя происходит сразу после волны возбуждения. Короткая задержка непосредственно перед этим восстановлением называется рефрактерным периодом. Первые 0,2 мс этой задержки называются абсолютным рефрактерным периодом, так как клетку не могут возбудить стимулы любой интенсивности. Следующий, более длинный период,— период относительной рефрактерности, так как для возникновения возбуждения необходимы импульсы большей, чем в норме, интенсивности. Он занимает около 1 мс после электрической деполяризации.
| Назад | Оглавление | Далее |