Медицинская реабилитация
1.6. Природа мышечного тонуса
Структурной основой деятельности поперечнополосатых мышц является удлиненная мышечная клетка (волокно). У млекопитающих и человека мышечные волокна вместе с рецепторными образованиями объединены в структурно-функциональные рефлекторные единицы — нервно-мышечные веретена (fusus). По отношению к местоположению в этих веретенах волокна поперечно-полосатых мышц делятся на экстрафузапьные и интрафузальные. Экстрафузальные волокна подразделяют на быстрые (фазические) и медленные (тонические). Быстрые, фазические, волокна составляют основу быстрых мышц, например разгибателей конечностей; медленные волокна — медленных, например камбаловидной и поясничной. Однако большинство мышц содержат как быстрые, так и медленные волокна. Тонические мышечные волокна могут оставаться в сокращенном состоянии в течение нескольких минут.
Мышечные волокна, обеспечивающие быструю фазную двигательную активность из-за отсутствия или незначительного содержания миоглобина, являются белыми. Мышечные волокна, способные к длительному неколебательному поддержанию сокращения в связи с высоким содержанием миоглобина, относят к красным. Белые мышечные волокна в гистохимической структуре имеют упорядоченные внутриклеточные мембраны и много миофибрилл при сравнительно небольшом объеме саркоплазмы, тогда как для красных характерно хаотичное расположение внутриклеточных мембран и обилие цитоплазматических органелл, в особенности митохондрий. Фазические волокна по размеру являются большими, имеют высокую концентрацию гликогена и энзимов, участвующих в анаэробном окислении. Тонические волокна по размеру меньшие и имеют высокую концентрацию энзимов, участвующих в аэробном окислении.
Различные по типу скелетные мышцы (медленные, или тонические, и быстрые, или фазические) иннервируются различными мотонейронами. Мотонейроны передних рогов спинного мозга в зависимости от размера разделяют на альфа-мотонейроны (более 25 мкм) и гамма-мотонейроны (менее 25 мкм). Различают два типа альфа-мотонейронов: альфа-большие — иннервируют мышечные волокна, способные совершать быстрые сокращения (фазические мотонейроны); альфа-малые — иннервируют мышечные волокна, которые относительно длительное время могут оставаться в сокращенном состоянии, что необходимо для поддержания мышечного тонуса и позы (тонические мотонейроны). Гамма-мотонейроны иннервируют интрафузальные мышечные волокна.
В основе рефлекторных механизмов, обеспечивающих выполнение двигательных функций и регуляцию мышечного тонуса, лежит простейший рефлекс на растяжение, или миотатический рефлекс. Рефлекс на растяжение имеет две формы (два компонента): медленный, или тонический, возникающий при медленном растяжении мышцы, и быстрый, или фазический, наблюдающийся при быстром растяжении. Мышечный тонус, исследуемый невропатологом при пассивных движениях конечности, относится к проявлениям тонического рефлекса на растяжение, а сухожильные рефлексы являются фазическими. Тонический компонент обусловливается возникновением афферентных импульсов, обеспечивающих продолжительный разряд мотонейронов, сопровождающийся длительным сокращением мышечных волокон, необходимым для под-, держания мышечного тонуса или соответствующей позы. В противоположность этому фазический компонент рефлекса возникает в результате кратковременного разряда афферентных импульсов и соответственно непродолжительного разряда мотонейронов, обусловливающего кратковременную стимуляцию мышечных волокон.
Рецептор миотатического рефлекса — мышечное веретено — состоит из нескольких тонких миниатюрных интрафузальных (внутри-веретенных) мышечных волокон, заключенных внутри капсулы. Эк-страфузальные (вневеретенные) мышечные волокна в 3—5 раз толще интрафузальных. Общее количество интрафузальных мышечных волокон колеблется от 2—3 до 10—15, а максимальная длина может достигать 1—1,5 см (рис. 8).
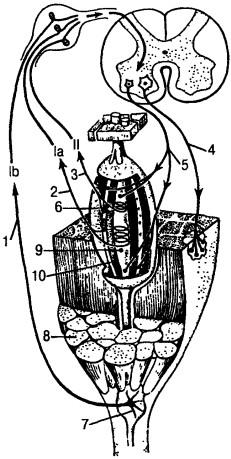
Рис. 8. Схема афферентных аппаратоа мышц:
1 — афферент lb, 2 — афферент 1а, 3 — афферент II, 4 — эфферент альфа-мотонейрона, 5 — эфферент гамма-мотонейрона, 6 — нервно-мышечное веретено, 7 — сухожильный рецептор, 8 — экстрафузальное мышечное волокно, 9—10 — интрафузальные мышечные волокна
Количество нервно-мышечных веретен в скелетной мышце тем больше, чем более тонкие и дифференцированные операции выполняет данная мышца. Так, в мышцах шеи, лица, предплечья количество веретен примерно в 15—20 раз больше, чем в мышцах бедра и таза.
Главная черта мышечного веретена — наличие толстого первичного афферентного нервного волокна, оканчивающегося в центральной области веретена в виде спирали, обвивающейся вокруг мышечного волокна. Ядра одних интрафузальных волокон веретена группируются в экваториальной области, образуя ядерную сумку, ядра других выстраиваются в ядерные цепи. Более тонкие чувствительные волокна, оканчивающиеся по обеим сторонам от центральной области, называются вторичными афферентами веретена. По обе стороны от центральной вязкоупругой части волокна отмечена типичная мышечная исчерченность. На этих мышечных частях рецептора оканчиваются тонкие двигательные волокна (гамма-аксоны; рис. 9).
При растяжении мышцы витки спирали мышечного рецептора растяжения (механорецептора), образованные безмиелиновой частью волокна, отходят друг от друга и в них возникает рецепторный потенциал — деполяризация (рис. 10).
Сухожильные органы Гольджи расположены в виде гроздьевидных нервных окончаний на сухожильных концах мышц и относятся к сенсорным образованиям (рецепторам).
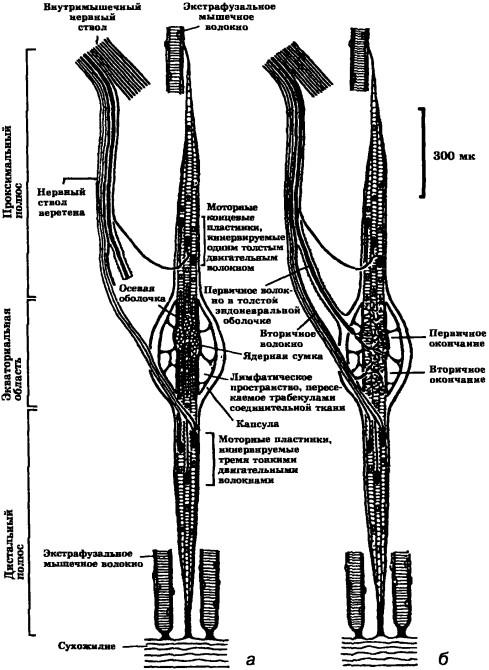
Рис. 9. Мышечные рецепторы млекопитающих (а) и схема их иннервации (б)
Веретена соединены с мышечными волокнами параллельно, сухожильные органы Гольджи — последовательно. Это означает, что при пассивном растяжении всей мышцы (например, если потянуть за сухожилие) следует ожидать возбуждения рецепторов обоих видов. Однако при активном сокращении мышцы ее веретена «разгружаются» (их натяжение ослабевает) и частота импульсации в соответствующих афферентах снижается. Наоборот, сухожильный орган, растягиваясь и сужаясь при активном сокращении, возбуждается.
Различие в действии активного мышечного сокращения на частоту импульсации этих двух видов рецепторов экспериментально показано Б. Мэттьюзом в 1933 г. Выделяя одиночные волокна из мышечных нервов, автор установил, что при активном сокращении мышцы в некоторых волокнах частота импульсации повышается (афференты сухожильных органов), а в других — понижается (афференты веретен; рис. 11).
Таким образом, в одиночном мышечном афферентном волокне, если мышца не растянута, регистрируется низкая частота разрядов. Если, раздражая вентральный корешок или непосредственно мышцу, вызвать ее сокращение, напряжение мышечного веретена во время укорочения снизится и частота разрядов в афферентах веретена уменьшится. Это уменьшение частоты импульсации при мышечном сокращении называется паузой и служит признаком афферента веретена. Напротив, если таким же образом выделенное из дорсального корешка волокно отходит от сухожильного органа, то сокращение приведет к увеличению частоты импульсации в его афферентах (рис. 12).
Сочетанная деятельность мышечных веретен и сухожильных рецепторов обеспечивает афферентную информацию о состоянии скелетных мышц, степени сокращения или растяжения мышечных волокон, натяжении сухожилий, о величине суставного угла и скорости движения в суставе.
Афферентные волокна мышечных веретен и сухожильных органов принадлежат к подгруппе I волокон группы А. Более толстые и быстро проводящие афференты принято относить к первичным волокнам веретена (подгруппа la), а более тонкие с более высоким порогом — к афферентам сухожильных органов Гольджи (подгруппа lb). Волокна подгруппы la и lb по-разному оканчиваются в спинном мозге. Волокна подгруппы la вызывают моносинаптический рефлекс: возбуждение (облегчение) мотонейронов мышц-синергистов и торможение мышц-антагонистов. Сухожильные органы Гольджи обеспечивают механизмы «обратного миотатического рефлекса». Они вызывают торможение мотонейрона своей мышцы и возбуждение мотонейрона мышцы-антагониста. В обоих случаях участвует по крайней мере один дополнительный синапс.
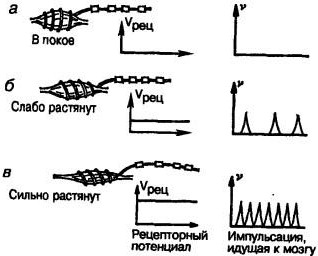
Рис. 10. Работа мышечного рецептора: а — рецепторное волокно в покое, рецепторный потенциал не возникает, и импульсы в нервную систему не поступают; б — волокно слабо растянуто, в «спирали* возникает рецепторный потенциал, в нервную систему идут импульсы; в — волокно сильно растянуто, величина рецепторного потенциала и частота импульсов, идущих к мозгу, возрастают
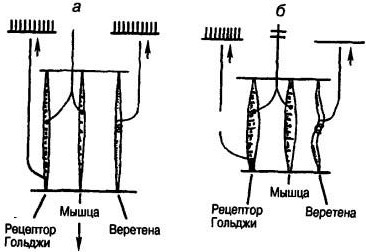
Рис. 11. Положение мышечных рецепторов относительно сокращающегося мышечного волокна. Сухожильные органы Гольджи соединены с мышечными волокнами последовательно, веретена — параллельно:
а — растяжение мышцы увеличивает частоту импульсации обоих рецепторов; б — активное сокращение мышцы усиливает импульсацию в сухожильном органе Гольджи и ослабляет разряд веретен
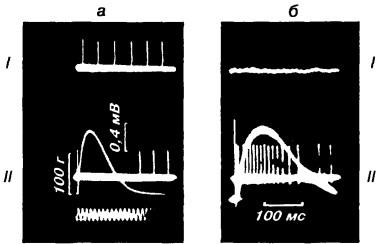
Рис. 12. Изменение частоты импульсации афферентов мышечных рецепторов в дорсальном корешке:
а — афферент веретена: I — фоновый уровень импульсации; II — импульсации того же афферента при сокращении мышцы уменьшается. Это снижение частоты импульсации называется паузой и свидетельствует о том, что исследуемое волокно представляет собой афферент веретена; б — афферент сухожильного органа Гольджи: I — при расслабленной мышце не импульсирует, II — при сокращенной — активируется
Вторичные афференты веретен относятся к подгруппе II. Качественно они реагируют на мышечное сокращение так же, как первичные афференты.
Кроме толстых афферентов в 1930 г. J. С. Eccles и С. S. Sherrington обнаружили тонкие волокна, попадающие в гамма-подгруппу (у) группы А (табл. 1). Установлено, что раздражение гамма-аксонов не приводит к появлению двигательного акта, но повышает частоту импульсации в афферентах веретен. Это объясняется тем, что гамма-активация сопровождается сокращением полярных отделов интрафузальных волокон и растяжением экваториальной части, тем самым изменяется исходная чувствительность рецепторов-спиралей к растяжению. Следовательно, при увеличении частоты раздражения гамма-волокон частота импульсации афферентов растет. Повышенная активность гамма-моторной системы может компенсировать уменьшение длины мышцы (рис. 13). Двигательные гамма-волокна влияют на импульсацию рецепторов веретен и не воздействуют на импуль-сацию сухожильных органов Гольджи.
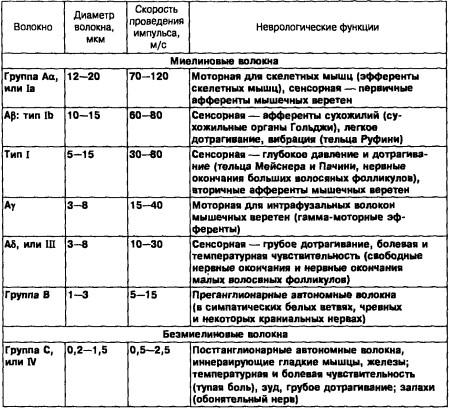
Таблица 1. Классификация типов нервных волокон в соответствии со скоростью проведения, диаметром и специфическими неврологическими функциями (Murray John, L Barr, A. Kiernan, 1993)
Таким образом, растяжение мышцы, веретена и интрафузаль-ных мышечных волокон вызывает формирование залпа афферентных импульсов к мотонейронам переднего рога спинного мозга и сокращение экстрафузальных волокон растянутой мышцы. Веретена-рецепторы постоянно находятся под влиянием небольшого растяжения, обусловленного силой тяжести. Идущие отсюда импульсы поступают в спинальные мотонейроны. Это приводит к их возбуждению и соответствующему сокращению экстрафузальных мышечных волокон, что дает возможность постоянно поддерживать мышечный тонус и необходимую позу. Благодаря системе вставочных нейронов спинного мозга реципрокная иннервация, открытая Н. Е. Введенским, вызывает расслабление антагонистов при каждом сокращении агонистов.
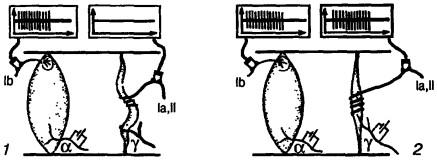
Рис. 13. Компенсирующее влияние повышенной активности гамма-моторной системы: 1 — мышца сокращена вследствие стимуляции альфа-мотонейронов, импульсация исходит от сухожильного рецептора, 2 — мышца сокращена, но одновременно стимулирован гамма-мотонейрон, импульсация исходит от афферентов веретена и сухожильного рецептора
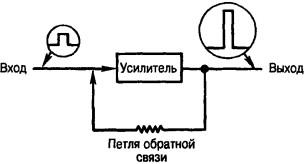
Рис. 14. Схема системы с обратной связью. Входной импульс напряжения усиливается и имеет на выходе большую амплитуду. Связь между выходом и входом (петлей обратной связи) может или увеличивать (положительная обратная связь), или уменьшать (отрицательная обратная связь) усиление
Первичные афференты веретен, альфа-мотонейроны и гамма-волокна образуют системы с обратной связью, аналогичные системам, встречающимся в технике (рис. 14). Часть выходного напряжения электронного усилителя отводится обратно на вход и образует петлю обратной связи. Если потенциал в петле обратной связи имеет тот же знак, что и входной потенциал, то это положительная обратная связь. Небольшое напряжение на входе усилителя увеличивается под действием усиленного напряжения, добавляемого через петлю обратной связи. Если потенциал в цепи обратной связи имеет противоположный знак по отношению к знаку входного потенциала, то выходное напряжение уменьшается, это пример отрицательной обратной связи. В технике отрицательная обратная связь применяется для увеличения стабильности системы за счет некоторого снижения коэффициента усиления.
Аналогично технической системе регуляция по типу обратной связи происходит в гамма-петле. Первичные афференты веретен можно считать входом, мышцу — усилителем, а связи и переключения в ЦНС — соединением между входом и усилителем. Гамма-волокна служат петлей обратной связи, действующей на вход системы.
Пример положительной обратной связи в гамма-петле: при рефлекторном раздражении гамма-мотонейроны (фузимоторные нейроны) получают возбуждающие импульсы, которые могут компенсировать веретено при укорочении мышцы, то есть гамма-активность усиливается вместе с усилением альфа-активности. Опыт показал, что альфа- и гамма-мотонейроны в одних случаях возбуждаются вместе, а в других — порознь, чего и следует ожидать при положительной обратной связи (С. Оке, 1969). Характер обратной связи (положительная или отрицательная) зависит от спинальных связей между сенсорными входами, фузимотор-ными клетками и альфа-мотонейронами. Отмечено, что афферентные волокна возбуждают фузимоторные клетки только через вставочные нейроны. Способность фузимоторных клеток отвечать на однократное раздражение ритмической импульсацией делает их больше похожими на вставочные нейроны, чем на мотонейроны. При исследование выявлено, что от 1/4 до 1/3 этих мотонейронов генерируют фоновые разряды; остальные без рефлекторной активности не функционируют. Ответ фузимоторных нейронов легче получить при раздражении нервов кожи, чем мышц. Ряд исследователей высказали предположение об управлении движениями через фузимоторные клетки. Однако возбудимость разных фузимоторных клеток, иннервирующих данную мышцу, сильно варьирует. Поэтому не исключено, что этот способ управления подходит только к небольшой группе наиболее возбудимых фузимоторных клеток.
При исследовании изменения возбудимости с помощью мо-носинаптического ответа во время мышечного напряжения установлено, что в течение первых 10 мс после начала сокращения возбудимость альфа-мотонейронов повышена, а затем следует продолжительный период сниженной возбудимости. Это торможение называется аутогенным, так как сокращение мышцы через активацию сухожильных рецепторов влияет на рефлекс, вызываемый в ее мотонейронном пуле (Р. Гранит, 1950). При введении новокаина в те области сухожилий, где локализуются рецепторы (сухожильные органы Гольджи), аутогенное торможение ослабляется.
В зависимости от уровня синаптической бомбардировки и фонового состояния ЦНС мотонейроны генерируют ритмические импульсы с частотой 10—50 в 1 с. В соответствии с различиями во времени сокращения медленных и быстрых мышц их мотонейроны делятся по свойствам. В ответ на тоническое растяжение медленная мышца (например, камбаловидная) развивает значительно большее напряжение, чем быстрая (например, икроножная), подвергнутая тому же воздействию. Частота импульсации мотонейронов медленной мышцы при тоническом рефлексе на растяжение составляет 15±5 в 1 с, а этого достаточно для достижения довольно слитного тетануса. Для быстрых мышц с их малым временем сокращения такая частота была бы недостаточной для слитного тетануса. Тоническое растяжение этих мышц приводит к разряду этих мотонейронов с большей частотой — до 40 (30— 60) в 1 с. Таким образом, частота импульсации двух типов мотонейронов согласуется с временем сокращения.
| Назад | Оглавление | Далее |