Медицинская реабилитация
8. Эффекторы
Сам термин эффектор позаимствован из кибернетики, а в физиологии эффекторами называют исполнительные или рабочие органы. К эффекторам относятся мышцы (скелетные, гладкие и сердечная) и железы внешней секреции. Скелетные мышцы называются также произвольными, поскольку их сокращением и расслаблением можно сознательно управлять, хотя частичное сокращение, обеспечивающее мышечный тонус, регулируется без участия сознания. При исследовании этих мышц путём световой микроскопии на продольных гистологических срезах можно видеть регулярную поперечную исчерченность, отчего мышцы получили название поперечнополосатых. Таким образом, термины скелетные, произвольные и поперечнополосатые являются синонимами и относятся к одной разновидности мышечной ткани. Управление этих мышц осуществляют мотонейроны, входящие в соматическую нервную систему.
Волокна сердечной мышцы тоже исчерчены поперечно, однако эта мышца не контролируется волей, а также имеет ряд морфологических и функциональных отличий от скелетных мышц. Сердечной мышцей управляют симпатические и блуждающие нервы, относящиеся к вегетативной нервной системе.
В гладких мышцах, расположенных в стенках крупных кровеносных сосудов и полых внутренних органов, нет поперечной исчерченности, Они почти не подчиняются сознательному контролю и управляются вегетативной нервной системой.
8.1. Строение скелетных мышц
Скелетные мышцы покрыты толстой соединительнотканной оболочкой. В толще мышцы менее толстая оболочка из соединительной ткани окружает как пучки мышечных волокон, так и отдельные волокна. Эта оболочка выполняет две функции. Во-первых, через неё к мышечным волокнам проходят нервы, кровеносные и лимфатические сосуды. Во-вторых, эластичная соединительная ткань объединяет тянущие усилия множества мышечных волокон.
Мышечные клетки или волокна имеют форму заострённых с концов цилиндров. Их тонкая цитоплазматическая мембрана называется сарколеммой (от греч. саркос – мясо, лемма – оболочка), а их цитоплазма – саркоплазмой. По периферии саркоплазмы располагаются многочисленные ядра мышечного волокна, а в центральной части – плотно уложены ориентированные продольно миофибриллы: в одном мышечном волокне их может содержаться больше тысячи.
Именно миофибриллы создают картину поперечной исчерченности мышечного волокна, в котором чередуются тёмные и светлые диски. Тёмные диски характеризуются двойным лучепреломлением в поляризованном свете, поэтому их называют анизотропными, в отличие от светлых изотропных дисков (Рис. 8.1). В середине каждого изотропного диска видна тёмная тонкая линия, получившая название Z-мембраны. Участок миофибриллы между соседними Z-мембранами называется саркомером, его длина в среднем составляет около 2-3 мкм.
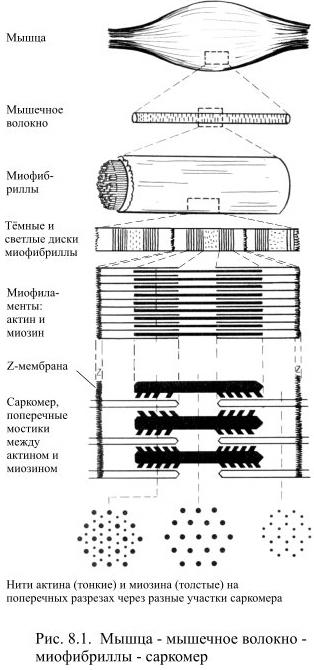
Саркомеры образованы нитями двух миофиламентов или сократительных белков – актина и миозина. Более тонкие нити актина одним концом прикрепляются к Z-мембране. К этой же мембране с другой стороны прикрепляются, как щетинки к щётке, актиновые нити соседнего саркомера. В промежутки между актиновыми нитями примерно на 1/4 их длины входят толстые нити второго сократительного белка – миозина. Участки миофибриллы, содержащие миозин соответствуют тёмным анизотропным дискам, а актин – светлым изотропным дискам. Мышечное сокращение осуществляется с помощью специального механизма втягивания тонких нитей актина в систему толстых нитей миозина. При сокращении уменьшается только длина саркомеров, тогда как длина актиновых и миозиновых нитей не изменяется.
Каждая тонкая нить образована двойной цепью глобулярных молекул мономеров актина, что можно сравнить с двойной ниткой бус, слегка скрученных по оси (Рис. 8.2).
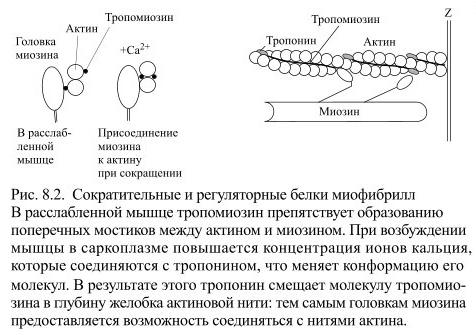
На каждой стороне такой двойной цепи есть продольные спиральные желобки. В желобках уложены длинные и тонкие молекулы регуляторного белка тропомиозина. К нитям тропомиозина присоединены молекулы второго регуляторного белка – тропонина, расположенные на одинаковом расстоянии друг от друга. Регуляторные белки контролируют соединения между нитями актина и миозина: от этого зависит сокращение и расслабление мышцы. Например, в расслабленной мышце молекулы тропомиозина занимают положение, препятствующее соединению миозиновых нитей с актиновыми.
У толстых нитей миозина есть многочисленные боковые ответвления, имеющие форму двойных головок. Эти головки могут поворачиваться, как на шарнирах, прикрепляться к нитям актина, а затем создавать тянущее усилие. Половина головок каждой нити миозина обращена в одну сторону, а половина – в другую, следовательно каждая половина головок может присоединиться к нитям актина, связанным как с одной, так и с другой Z-мембранами саркомера.
От сарколеммы мышечного волокна отходят внутрь узкие канальцы, получившие название поперечных Т-трубочек (Рис. 8.3).
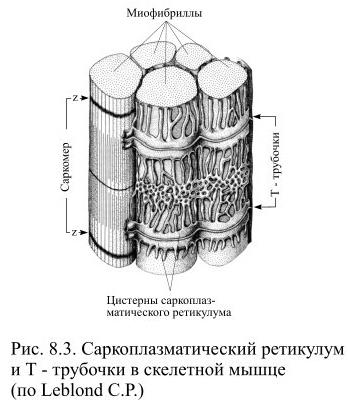
Когда возбуждение передаётся через нервно-мышечный синапс на сарколемму, волна деполяризации распространяется и на Т-трубочки. В саркоплазме с Т-трубочками соседствуют цистерны саркоплазматического ретикулума, представляющего мышечный аналог гладкого эндоплазматического ретикулума. Вокруг одного саркомера проходят две Т-трубочки, а к каждой из них с обеих сторон прилежат цистерны саркоплазматического ретикулума. Каждая Т-трубочка с двумя прилежащими к ней цистернами саркоплазматического ретикулума образует триаду. Цистерны ретикулума соединены друг с другом посредством множества продольных трубочек, образуя единую систему. В саркоплазматическом ретикулуме накапливаются ионы кальция, поступающие из саркоплазмы активным транспортом. Главный интегральный белок мембраны саркоплазматического ретикулума выполняет функцию кальциевого насоса и при расслаблении мышцы перекачивает ионы кальция в ретикулум, используя для этого энергию АТФ.
8.2. Механизм сокращения мышечных волокон
К мышечному сокращению приводит повышение концентрации ионов кальция в саркоплазме и их присоединение к тропонину. Вследствие этого молекула тропонина изменяет свою конформацию и потому сдвигает тропомиозин в глубину желобка актиновой нити. Так на актиновой нити освобождаются участки, служащие местом прикрепления головок миозина. Внутриклеточный кальций хранится в саркоплазматическом ретикулуме, представляющем систему продольных трубочек и поперечных цистерн. Когда возбуждение передастся через нервно-мышечный синапс на сарколемму, потенциалы действия распространяются по ней к Т-трубочкам, а в связи с этим деполяризуется соприкасающаяся с ними мембрана цистерн саркоплазматического ретикулума. Это приводит к открытию в ней каналов для ионов кальция. Кальций выходит по концентрационному градиенту и присоединяется к тропонину. Весь механизм преобразования процесса возбуждения в мышечное сокращение называется электромеханическим сопряжением. Ключевая роль в нём принадлежит ионам кальция.
Сокращение волокна происходит благодаря скольжению актиновых нитей вдоль миозиновых – этому способствует образование поперечных мостиков между ними. Длина актиновых и миозиновых нитей при этом не изменяется, но вследствие скольжения уменьшается длина саркомеров, а значит и всего мышечного волокна (Рис 8.4).
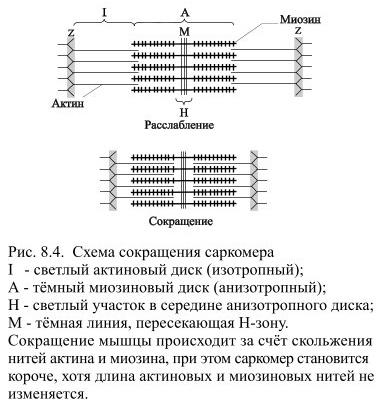
В образовании поперечных мостиков участвуют головки миозина, периодически прикрепляющиеся к соседним нитям актина и создающие усилие, подтягивающее нити актина. Поскольку миозиновые головки одновременно прикрепляются к актиновым нитям, соединённым с двумя соседними Z-мембранами, тянущее усилие сближает Z-мембраны, уменьшая тем самым длину саркомера.
Многочисленные головки миозина прикрепляются к актиновым нитям поочерёдно, а затем, после осуществления тянущего усилия – "гребка", отделяются от этого участка актина, чтобы тотчас прикрепиться к следующему. Эту деятельность можно сравнить с вытягиванием верёвки, которую человек перебирает обеими руками. Для ритмичных прикреплений и отделений миозиновых головок расходуется энергия АТФ, молекулы которой расщепляются под действием каталитически активных центров миозина и при участии активирующего этот процесс актина. Вероятно молекула АТФ связывается с поперечным мостиком после завершения "гребка", затем расщепляется, а освободившаяся энергия идёт на отделение миозиновой головки, которая тотчас же прикрепляется к соседнему участку актина. Существуют основания полагать, что при мышечном сокращении за одну секунду может происходить от 50 до 100 "гребков.
Высокая концентрация ионов кальция в саркоплазме активирует кальциевый насос, закачивающий эти ионы обратно в саркоплазматический ретикулум. При уменьшении концентрации кальция в саркоплазме тропонин освобождается от него и восстанавливает прежнюю конформацию молекулы. В связи с этим возвращается на прежнее место тропомиозин и уже не позволяет большинству головок миозина прикрепляться к актиновой нити. Когда большинство головок миозина начнёт отделяться от актиновых нитей, наступает расслабление мышечного волокна. Оно происходит пассивно, без затраты энергии: нити актина и миозина скользят друг относительно друга так, что саркомеры увеличивают свою длину.
8.3. Двигательные единицы
Любое мышечное волокно иннервируется только от одного мотонейрона. В то же время один мотонейрон иннервирует группу мышечных волокон, вместе с которыми он образует двигательную единицу. Она является наименьшей функциональной единицей моторной системы. В разных мышцах двигательные единицы сильно отличаются по количеству мышечных волокон. Так, например, в глазодвигательных мышцах на один нейрон приходится около десятка мышечных волокон, в двигательных единицах мелких мышц руки их около сотни, а в крупных мышцах конечностей и туловища – более тысячи на один мотонейрон. Малые единицы характерны для мышц, с помощью которых совершаются точные, хорошо координированные движения, тогда как большие преобладают в массивных мышцах, таких например, как мышцы спины.
В зависимости от скорости развития максимальной силы сокращения, а также от быстроты утомления различают три типа двигательных единиц: 1) быстрый, утомляемый тип; 2) медленный, мало утомляемый и 3) быстрый, мало утомляемый – этот тип является промежуточным: мышечные волокна сокращаются немного медленней, чем 1), а устают почти так же мало, как 2). В большинстве мышц представлены все три типа двигательных единиц, но в разном соотношении, которое варьирует от одного человека к другому. В спортивной физиологии людей с относительно большим процентом быстрых волокон относят к спринтерам, а обладателей большего, чем в среднем, процента медленных волокон – к стайерам. У всех людей при старении процент быстрых волокон уменьшается.
Волокна быстрых двигательных единиц относительно меньше снабжаются кровью и поэтому имеют бледно-красный цвет. Они предназначены для мощных быстрых сокращений в течение короткого времени, поскольку быстро утомляются, не поспевая из-за относительно малого кровоснабжения окислять глюкозу до конечных продуктов. Волокна медленного типа обеспечены обильной сетью кровеносных капилляров, что придаёт им тёмно-красный цвет. И, хотя они сокращаются медленнее, зато способны очень долго работать без утомления. Развиваемая ими сила составляет от 1 до 10% от силы быстрых волокон.
Тела мотонейронов двигательных единиц медленного типа имеют наименьшие размеры и самый низкий порог возбуждения, поэтому только они могут активироваться слабыми входными сигналами. Мотонейроны быстрых двигательных единиц, напротив, являются самыми крупными и, чтобы возбудить их, нужны самые сильные стимулы. Поэтому медленные двигательные единицы используются при любых движениях, а быстрые – только в тех случаях, когда понадобится большая сила сокращающихся мышц. Так, например, при стоянии или спокойной ходьбе сокращение мышц обеспечивает менее, чем половина всех двигательных единиц – здесь сила мышечного сокращения составляет около 20% от максимальной. Для того, что выполнить прыжок, может потребоваться максимальная сила: в таком случае понадобится активация мотонейронов быстрых двигательных единиц.
Таким образом, сила мышечных сокращений может регулироваться количеством активированных двигательных единиц. Такой механизм регуляции называется рекрутированием.
8.4. Зависимость мышечного сокращения от частоты нервных импульсов
Сила и скорость мышечного сокращения пропорциональны также частоте потенциалов действия, распространяющихся к мышце по аксону мотонейрона – это ещё один механизм регуляции, который называется частотным кодированием. Потенциал действия в нерве или мышце длится приблизительно 1-3 мс, а самое короткое сокращение, состоящее из укорочения и расслабления, продолжается примерно 10-100 мс (Рис 8.5). Таким сокращением мышца отвечает на единичный потенциал действия – это сокращение называется одиночным.
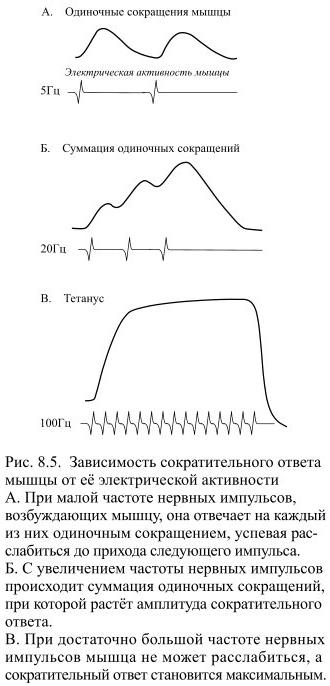
Если частота возбуждающих мышцу нервных импульсов станет расти, она не успеет полностью расслабиться к моменту действия очередного потенциала действия и сила её сокращения будет увеличиваться. Такой режим сокращения получил название зубчатого тетануса. С ещё большим увеличением частоты стимуляции формируется плато: в это время мышца совсем не расслабляется, а сокращение делается максимальным – этот режим называется гладким тетанусом. Серия следующих друг за другом потенциалов действия приводит к значительному повышению внутриклеточной концентрации кальция, поэтому сократительный ответ и становится сильнее и продолжительнее.
Частота поступающих к мышцам потенциалов действия варьирует в небольших пределах. Например, при произвольных сокращениях потенциалы действия в мотонейронах появляются с частотой приблизительно в 8 Гц, в редких случаях она может превысить 25 Гц. С нарастанием частоты поступающих к мышце потенциалов действия сила её сокращения увеличивается.
8.5. Режимы мышечных сокращений
Взаимодействие человека с окружающей средой невозможно представить без сокращений его мышц. Вполне определённо высказывался по этому поводу И. М. Сеченов: "Всё бесконечное разнообразие внешних проявлений мозговой деятельности сводится окончательно к одному лишь явлению – мышечному движению. Смеётся ли ребёнок при виде игрушки, улыбается ли Гарибальди, когда его гонят за излишнюю любовь к родине, дрожит ли девушка при первой мысли о любви, создаёт ли Ньютон мировые законы и пишет их на бумаге – везде окончательным фактом является мышечное движение".
Скелетные мышцы прикрепляются к костям и при своём сокращении изменяют положение какого-либо сустава. Мышцы, производящие движение в одном направлении, называются агонистами или синергистами, а обеспечивающие движение в противоположном направлении – антагонистами. Например, все мышцы, сокращение которых приводит к сгибанию конечности в суставе, будут синергистами, а мышцы, обеспечивающие разгибание, являются для них антагонистами.
При сокращении мышцы в ней могут изменяться два параметра: длина и мышечное напряжение. В зависимости от того, какое из этих изменений преобладает, различают два режима мышечных сокращений: изотонический и изометрический. В изотоническом режиме мышцы работают, перемещая какой-либо постоянный груз, как это происходит, например, при сгибании руки в локтевом суставе. В ней может находиться какой-нибудь предмет, хотя это и не обязательно, поскольку и сама рука является "грузом". Так как груз постоянен, постоянно и мышечное напряжение (изотонично), тогда как длина мышцы изменяется (в приведённом примере уменьшается длина сгибателей).
В качестве примера работы мышц в изометрическом режиме можно рассмотреть попытку поднять, например, ведро с водой, Для того, чтобы оторвать его от пола, мышцы должны наращивать напряжение, длина же их остаётся постоянной (изометричной) до тех пор, пока не изменится угловое положение сустава. После того, как груз придёт в движение, мышцы переходят в изотонический режим работы. Если зафиксировано постоянное угловое положение сустава (поэтому длина мышц не изменяется) и происходит мышечное сокращение (для того, например, чтобы поднять или сдвинуть тяжёлый неподвижный предмет или удерживать какой-нибудь груз), то режим работы является изометрическим.
Если при сокращении длина мышцы уменьшается в связи с увеличением развиваемой силы, такой режим называют ауксотоническим. Он является переходным между изотоническим и изометрическим режимами, поскольку включает две существующие фазы сокращения – динамическую и статическую.
8.6. Регуляция длины и напряжения мышц
Сокращения мышц контролируются двумя видами проприоцепторов: интрафузальным рецепторами и сухожильными рецепторами Гольджи (См. рис. 7.2). Мышечное веретено представляет собой соединительнотканную капсулу, которая окружает группу особых, интрафузальных (лат. fusus – веретено) мышечных волокон. Они тоньше и короче обычных экстрафузальных волокон: всё веретено имеет длину от 4 до 10 мм. В средней части интрафузального волокна сократительных белков нет, там находятся окончания чувствительного нейрона, которые возбуждаются при растяжении волокна. На обоих концах интрафузального волокна есть сократительные белки, они иннервируются особой разновидностью мотонейронов, которые называются g-мотонейронами в отличие от a-мотонейронов, предназначенных лишь для экстрафузальных волокон.
Когда внешняя сила растягивает мышцу, возбуждение от рецепторов интрафузальных волокон увеличивается (См. рис. 7.3). Чувствительный нейрон двумя коллатералями своего аксона одновременно передаёт возбуждение a- и g-мотонейронам и вслед за этим происходит одновременное возбуждение, а затем и сокращение интра- и экстрафузальных волокон: это описанный в главе 7 рефлекс растяжения, который обеспечивает регуляцию длины мышцы.
Центральная нервная систему с помощью g-мотонейронов контролирует чувствительность мышечных веретён. Так, например, g-мотонейроны могут возбуждаться двигательными ядрами головного мозга, что приведёт к сокращению концевых участков интрафузальных волокон и растяжению центральной их части, в которой находятся окончания чувствительного нейрона. Это вызовет возбуждение чувствительного нейрона, он подействует на a- и g-мотонейроны, что закончится небольшим сокращением мышцы – повышением её тонуса.
Таким образом, мышечный тонус регулируется с помощью g-мотонейронов. Если двигательные ядра головного мозга повысят их активность, то и мышечный тонус повысится. Но, если влияние головного мозга на g-мотонейроны будет опосредовано тормозными интернейронами спинного мозга, то их активность, а затем и мышечный тонус уменьшатся. В физиологических условиях часто бывает необходимо повышать тонус одних мышц и понижать – тонус других, поэтому происходит и стимуляция, и торможение активности различных g-мотонейронов. Благодаря этому одни мышцы могут находиться в большем тонусе, чем другие – именно это позволяет удерживать определённую позу и переходить от неё к движению. Состояние тревоги, неуверенности, страха обычно сопровождается увеличением тонуса всех мышц, механизм этого явления состоит в усилении активирующих и уменьшении тормозных влияний головного мозга на g-мотонейроны.
Сухожильные рецепторы Гольджи почти не реагируют на изменение длины мышцы, но возбуждаются при изменении мышечного напряжения. Чем оно сильнее, тем больше натягиваются коллагеновые волокна сухожилий, прикрепляющие мышцу к костям. Рецепторы Гольджи, вплетённые в ткань сухожильных волокон, при этом сдавливаются, а распространяющееся от них по чувствительному нейрону возбуждение переключается в спинном мозгу на тормозной интернейрон (См. рис. 7.5), уменьшающий активность мотонейрона и ограничивающий тем самым дальнейший рост мышечного напряжения.
8.7. Гладкие мышцы
Большинство гладких мышц располагается в стенках кровеносных сосудов и полых внутренних органов: это желудок и кишечник, выносящие протоки пищеварительных желёз, мелкие бронхи, мочевой пузырь и мочеточники и т.д. В типичном варианте они образуют два мышечных слоя: внутренний циркулярный и наружный продольный. Актин и миозин в гладких мышцах расположены не так упорядоченно, как в скелетных, и поэтому в них нет поперечнополосатой исчерченности. По сравнению со скелетными гладкие мышцы расходуют меньше энергии, они лучше приспособлены для продолжительных сокращений и очень медленно утомляются. Их одиночные сокращения продолжительны, они могут длиться несколько секунд, а тетанус в гладких мышцах возникает при малой частоте стимуляции – около 1,5-2 Гц.
Гладкие мышцы иннервирует вегетативная нервная система, деятельность которой не подчинена сознательному контролю и является непроизвольной. Контроль за сокращениями гладких мышц в большинстве случаев осуществляют оба отдела вегетативной нервной системы – симпатический и парасимпатический, причём характер их влияний на сокращения гладких мышц обычно противоположен. Кроме того, деятельность гладких мышц регулируют гуморальные и местные механизмы.
У некоторых гладких мышц, например, управляющих шириной зрачка или сокращениями мускулатуры семявыносящего протока, отдельно иннервируется каждое волокно, что позволяет им сокращаться относительно быстро. Но у большинства гладких мышц лишь немногие волокна образуют синапсы непосредственно с вегетативными нейронами. Вслед за возбуждением таких волокон сигналы к соседним с ними передаются электротонически – через щелевые контакты или электрические синапсы.
Гладкие мышцы способны сами спонтанно генерировать возбуждение – это их свойство называется автоматией. Даже при отсутствии каких-либо нервных или гуморальных влияний некоторые волокна (их называют водителями ритма или пейсмекерами) периодически деполяризуются до критического уровня, а возникшее возбуждение распространяется через щелевые контакты на соседние клетки. После этого гладкие мышцы медленно сокращаются.
Характер влияния вегетативных нейронов на гладкие мышцы определяется типом нейромедиаторов – ацетилхолина или норадреналина и постсинаптических рецепторов (М-холинорецепторы, a- и b-адренорецепторы). Вегетативные нейроны не инициируют возбуждение и сокращение гладких мышц: они регулируют частоту и длительность этих непрерывно происходящих процессов.
Гладкие мышцы отличаются пластичностью – способностью приспосабливаться к растяжению, а не отвечать на него, как скелетные мышцы, немедленным сокращением. Например, при наполнении мочевого пузыря его стенки растягиваются без ответного сокращения мышц и за счёт этого давление в пузыре повышается не сильно.
Медленные ритмические сокращения гладких мышц желудочно-кишечного тракта постоянно перемешивают съеденную пищу с пищеварительными соками. Особые формы сокращений – перистальтические волны продвигают её, мощные циркулярные слои волокон – мышечные жомы регулируют это продвижение, а также выделение в просвет кишечника пищеварительных секретов поджелудочной железы и печени. Тонус гладких мышц сосудов поддерживает необходимую величину кровяного давления. Длительное тоническое сокращение гладких мышц не приводит, в отличие от скелетных, к быстрому развитию утомления, что связано с особенностями энергетического обмена.
8.8. Сердечная мышца – миокард
Миокард состоит из отдельных поперечнополосатых клеток, прочно соединённых конец в конец так, что образуется нитевидная цепочка, которую часто называют волокном. Миофибриллы сердечной мышцы не обособлены, как в скелетной, и часто пересекаются друг с другом, образуя непрерывную общую сеть. Саркоплазматический ретикулум развит меньше, чем в скелетной мышце, но связанные с Т-трубочками узкие переплетающиеся цистерны прилегают к поверхности каждого саркомера.
Потенциал покоя у клеток миокарда составляет около -90 мВ, а потенциал действия длится примерно 0,3 секунды (Рис 8.6), что гораздо больше, чем в скелетной мышце, где его продолжительность составляет лишь несколько тысячных долей секунды. Благодаря этому все сокращения сердечной мышцы являются одиночными и тетанус в ней невозможен.: сокращение длится примерно столько же времени, что и возбуждение. Мышца при этом рефрактерна, т.е. не может ответить ни на какой стимул.
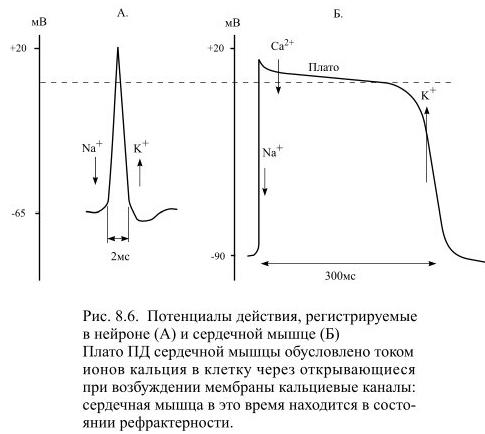
Столь необычная длительность возбуждения сердечной мышцы обусловлена тем, что в образовании потенциала действия помимо натриевых каналов дополнительно участвуют каналы для ионов кальция: ток кальция увеличивает время деполяризации, образуя т.н. плато потенциала действия.
Отдельная группа особых волокон миокарда образует в нём проводящую систему сердца (Рис. 8.7).
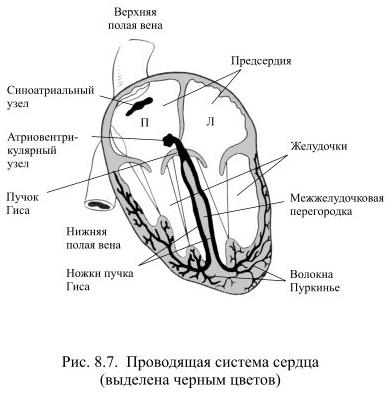
Она состоит из синусно-предсердного узла (или синоатриального), расположенного в стенке правого предсердия, предсердно-желудочкового узла (или атриовентрикулярного), находящегося в основании межпредсердной перегородки: от этого узла отходит пучок Гиса, который в межжелудочковой перегородке разделяется на две ножки, заканчивающиеся многочисленными волокнами Пуркинье в миокарде обоих желудочков.
Синоатриальный узел – это генератор возбуждения в миокарде, его главный пейсмекер, водитель ритма. Он образован клетками, мембрана которых даже в покое пропускает небольшое количество ионов натрия. Вход натрия деполяризует мембрану до критического уровня и тогда возникает потенциал действия, быстро распространяющийся по проводящей системе на волокна рабочего миокарда. Таким образом, возникновение возбуждения в миокарде вызывается спонтанной деполяризацией водителя ритма, т.е. происходит автоматически.
Собственный ритм сердца можно определить только при полной вегетативной блокаде, т.е. при исключении влияний блуждающих (парасимпатических) и симпатических нервов. Можно вычислить собственный ритм сердца по формуле: 118,1 – (0,57 ´ возраст), что в двадцатилетнем возрасте составит приблизительно 107 сокращений в минуту.
У здоровых людей в этом возрасте частота сокращений сердца составляет приблизительно 75 в минуту, что объясняется постоянным влиянием блуждающего нерва, который урежает частоту сокращений сердца. Напротив, симпатические нервы могут, при повышении своей активности увеличивать частоту и силу сердечных сокращений.
Вегетативные нервы сердца находятся в постоянном тонусе, т.е. проводят к нему из центральной нервной системы несколько нервных импульсов в секунду. В покое преобладает тонус блуждающего нерва, чем и объясняется меньшая, по сравнению с собственным ритмом сердца, частота сердечных сокращений. При физической работе или эмоциональных переживаниях повышается тонус симпатических нервов, что вызывает увеличение частоты сокращений сердца. Тонус блуждающих и симпатических нервов может изменяться рефлекторно.
8.9. Железы
Железы внешней секреции или экзокринные имеют трубчатые протоки для выведения образующегося в них секрета. Секрет может выводиться наружу, как у потовых или у сальных желёз, он может поступать в ротовую полость, как у слюнных желёз, в кишечник, как у печени и поджелудочной железы.
Секреторная активность желёз регулируется нервными, гуморальными и местными механизмами, причём значимость каждого из механизмов для деятельности разных желёз неодинакова. Нервный контроль за самой секрецией и выведением секрета осуществляет вегетативная нервная система, не находящаяся под произвольным контролем. Секреторная деятельность большинства пищеварительных желёз усиливается под влиянием парасимпатического отдела. Повышение тонуса парасимпатических нервов происходит рефлекторно: в естественных условиях это вызывается актом еды и поступлением пищи в желудочно-кишечный тракт.
Резюме
Скелетные мышцы контролируются исключительно мотонейронами, от которых они получают возбуждение. Возбуждение мышц приводит к выходу кальция из саркоплазматического ретикулума и присоединению его к регуляторному белку, что даёт возможность сократительным белкам взаимодействовать. Сокращение мышцы обеспечивается скольжением нитей миозина вдоль актиновых нитей. Контроль длины и напряжения мышц осуществляется с помощью двух типов проприоцепторов. Управление активностью g-мотонейронов позволяет регулировать мышечный тонус. Гладкие мышцы и сердечная мышца обладают свойством автоматии – они способны сами генерировать возбуждение. Их деятельность контролируется вегетативной нервной системой.
Вопросы для самоконтроля
115. Как называется цитоплазматическая мембрана мышечного волокна?
А. Саркоплазма; Б. Сарколемма; В. Саркоплазматический ретикулум; Г. Саркомер; Д. Миофибрилла.
116. Нити какого белка в миофибрилле прикреплены к Z-мембране?
А. Тропонина; Б. Тропомиозина; В. Актина; Г. Миозина; Д. Эластина.
117. Какие белки участвуют в образовании поперечных мостиков при сокращении мышечного волокна?
А. Тропонин – тропомиозин; Б. Тропонин- актин; В. Тропомиозин – актин; Г. Миозин – актин; Д. Миозин – тропонин.
118. Как называется участок миофибриллы между соседними Z-мембранами?
А. Актиновый диск; Б. Миозиновый диск; В. Триада; Г. Саркоплазма; Д. Саркомер.
119. Какой из перечисленных ниже белков является сократительным?
А. Миозин; Б. Тропомиозин; В. Тропонин; Г. Саркомер; Д. Сарколемма.
120. Что из перечисленного ниже в мышечном сокращении выполняет роль регуляторного белка?
А. Актин; Б. Тропомиозин; В. Миозин; Г. Миофиламент; Д. Саркомер.
121. Как называется мотонейрон с иннервируемыми им мышечными волокнами?
А. Нервно-мышечный синапс; Б. Триада; В. Двигательная единица; Г. Миофибрилла; Д. Эффектор.
122. Какой процесс приводит к сокращению мышечного волокна?
А. Сокращение нитей актина; Б. Сокращение нитей миозина; В. Совместное сокращение нитей миозина и актина; Г. Скольжение нитей актина и миозина относительно друг друга; Д. Для сокращения необходимы все указанные процессы.
123. Какое событие при мышечном сокращении должно произойти раньше остальных?
А. Присоединение ионов кальция к молекулам тропонина; Б. Повышение концентрации ионов кальция в межфибриллярном пространстве; В. Перемещение тропомиозина в глубину желобка между нитями актина; Г. Прикрепление миозиновых головок к актину; Д. Расщепление молекул АТФ под влиянием каталитических центров миозина.
124. Для какого процесса используется энергия АТФ, освобождающаяся под влиянием миозина?
А. Выход ионов кальция из цистерн саркоплазматического ретикулума; Б. Присоединение ионов кальция к тропонину; В. Перемещение молекулы тропомиозина в глубину желобка актиновой нити; Г. Разъединение поперечных мостиков; Д. Повышение концентрации ионов кальция в межфибриллярном пространстве.
125. Что происходит сразу после возбуждения поперечных Т-трубочек мышечного волокна?
А. Выход ионов кальция из саркоплазматического ретикулума; Б. Возврат кальция в цистерны саркоплазматического ретикулума; В. Присоединение кальция к тропонину с изменением его конформации; Г. Соединение кальция с тропонином без последующего изменения конформации; Д. Расщепление АТФ на актиновых нитях.
126. Что произойдёт, если на концевой пластинке, принадлежащей мышечному волокну, возникнет подпороговый постсинаптический потенциал?
А. Одиночное сокращение; Б. Зубчатый тетанус; В. Гладкий тетанус; Г. Все ответы верны; Д. Все ответы не верны.
127. Что произойдёт, если сокращающееся мышечное волокно будет возбуждаться разрядами мотонейронов, следующими до начала фазы расслабления?
А. Суммация одиночных сокращений; Б. Зубчатый тетанус; В. Гладкий тетанус; Г. Никаких изменений не произойдёт в связи с рефрактерностью волокна; Д. Все ответы не верны.
128. Какое из перечисленных ниже свойств имеется у гладких мышц и отсутствует у скелетных?
А. Возбудимость; Б. Проводимость; В. Сократимость; Г. Автоматия; Д. Лабильность.
129. У какой из указанных мышц следует предполагать наибольшую длительность потенциала действия?
А. Скелетная мышца с высоким процентом быстрых двигательных единиц; Б. Скелетная мышца с преобладанием медленных двигательных единиц; В. Скелетная мышца промежуточного типа; Г. Гладкая мышца; Д. Сердечная мышца.
| Назад | Оглавление | Далее |